“𝐓𝐡𝐞 𝐪𝐮𝐞𝐬𝐭𝐢𝐨𝐧 𝐢𝐬 𝐧𝐨𝐭 𝐰𝐡𝐚𝐭 𝐲𝐨𝐮 𝐥𝐨𝐨𝐤 𝐚𝐭, 𝐛𝐮𝐭 𝐰𝐡𝐚𝐭 𝐲𝐨𝐮 𝐬𝐞𝐞.” — 𝐇𝐞𝐧𝐫𝐲 𝐃𝐚𝐯𝐢𝐝 𝐓𝐡𝐨𝐫𝐞𝐚𝐮 For decades, Alzheimer’s research has looked at the same data: amyloid plaques, tau tangles, neuroinflammation. But what if we have been seeing it wrong?
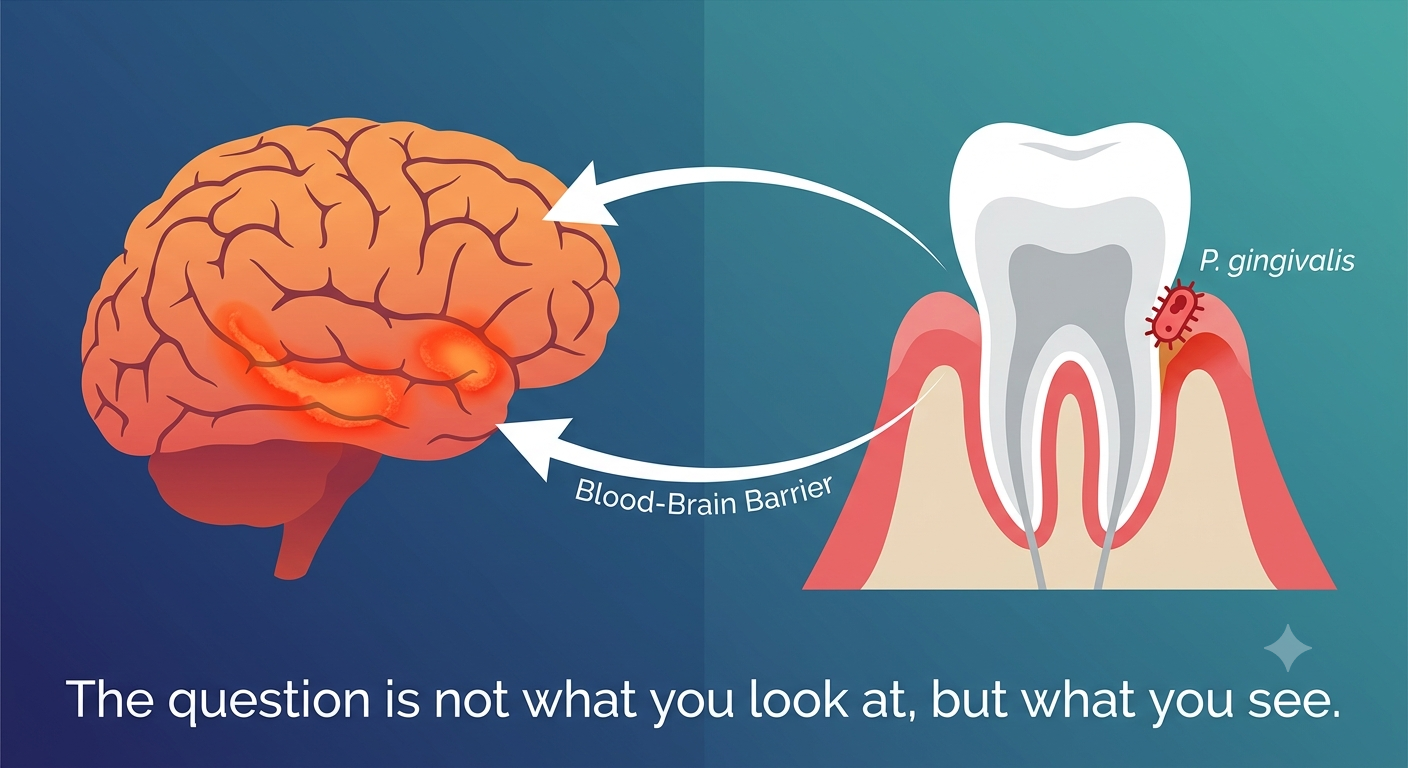
Two underappreciated forces—chronic stress and an oral bacterium called P. gingivalis—may be converging on the same vulnerable brain circuits, accelerating neurodegeneration in ways that neither factor alone can explain.
This “dual-hit” model reframes Alzheimer’s not as a single pathological cascade, but as an interaction between host vulnerability and environmental exposures. It asks a different question: not how do we treat late-stage dementia? but why do some brains age better than others?
Cellular Senescence: Persistent exposure to P. gingivalis toxins promotes the early accumulation of senescent (aging, non-dividing) cells in structural tissues like bone and oral mucosa, accelerating local tissue breakdown and bone resorption.
Are you looking at someone with Alzheimer’s disease? Or do you see beyond the disease to possible causes?
Metabolic Disruption: P. gingivalis alters the gut microbiome and systemic oxidative pathways, which is strongly associated with increased insulin resistance and elevated risk for metabolic complications during aging.
If you’d like, I can elaborate on:
Specific oral hygiene and nutritional strategies to lower P. gingivalis risk.
The connection between periodontal health and cardiovascular aging.
#AlzheimersDisease #Neuroscience #HealthyAging #BrainHealth #Stress #OralHealth #PorphyromonasGingivalis #Neuroinflammation #CognitiveResilience
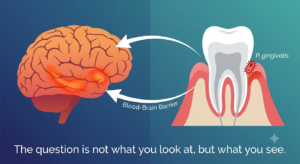
𝐈𝐧 𝐭𝐡𝐞 𝐰𝐨𝐫𝐥𝐝 𝐨𝐟 𝐛𝐢𝐨𝐦𝐞𝐝𝐢𝐜𝐚𝐥 𝐫𝐞𝐬𝐞𝐚𝐫𝐜𝐡, 𝐰𝐞 𝐟𝐫𝐞𝐪𝐮𝐞𝐧𝐭𝐥𝐲 𝐬𝐩𝐞𝐧𝐝 𝐝𝐞𝐜𝐚𝐝𝐞𝐬 𝐬𝐭𝐚𝐫𝐢𝐧𝐠 𝐝𝐢𝐫𝐞𝐜𝐭𝐥𝐲 𝐚𝐭 𝐞𝐱𝐩𝐚𝐧𝐬𝐢𝐯𝐞 𝐜𝐥𝐢𝐧𝐢𝐜𝐚𝐥 𝐝𝐚𝐭𝐚𝐬𝐞𝐭𝐬 𝐰𝐡𝐢𝐥𝐞 𝐜𝐨𝐦𝐩𝐥𝐞𝐭𝐞𝐥𝐲 𝐦𝐢𝐬𝐬𝐢𝐧𝐠 𝐭𝐡𝐞 𝐬𝐭𝐫𝐮𝐜𝐭𝐮𝐫𝐚𝐥 𝐜𝐨𝐧𝐧𝐞𝐜𝐭𝐢𝐨𝐧𝐬 𝐬𝐭𝐚𝐫𝐢𝐧𝐠 𝐛𝐚𝐜𝐤 𝐚𝐭 𝐮𝐬. When scientific fields remain tightly siloed—treating systemic health, chronic stress, and neurodegeneration as entirely separate universes—we risk treating isolated symptoms while missing the integrated biological reality. “The question is not what you look at, but what you see.” ― Henry David Thoreau
Real breakthroughs rarely emerge from finding entirely new data points in the laboratory; instead, they come from fundamentally changing how we look at the data we already possess. By refusing to cross traditional disciplinary boundaries, researchers trap themselves in legacy frameworks that obscure how peripheral and central systems interact over the human lifespan. True scientific progress requires stepping back from specialized myopia to observe the broader environmental and physiological pressures acting on biological systems.
This institutional inertia often blinds investigators to the true nature of chronic multi-system pathologies that do not fit neatly into a single medical specialty. When complex conditions are viewed through a single narrow lens, the cumulative impact of overlapping risk factors goes entirely unrecognized by traditional diagnostic models. Expanding our analytical perspective allows us to perceive the subtle, multi-hit trajectories that precede clinical disease onset.
Challenging these entrenched silos is essential if we want to move beyond stagnant treatment paradigms and address the root causes of cognitive aging. By welcoming interdisciplinary frameworks, the scientific community can begin to bridge the artificial gaps separating immunology, endocrinology, and neurology. This broader perspective ultimately revitalizes how we evaluate clinical evidence and design preventive interventions.
Looking forward to sharing more on how cross-disciplinary blind spots shape our understanding of cognitive aging soon. These ongoing inquiries highlight the urgent need for a more unified approach to translational research and clinical trial design. Engaging with these deeper conceptual challenges will help redefine the future landscape of preventive health strategy.




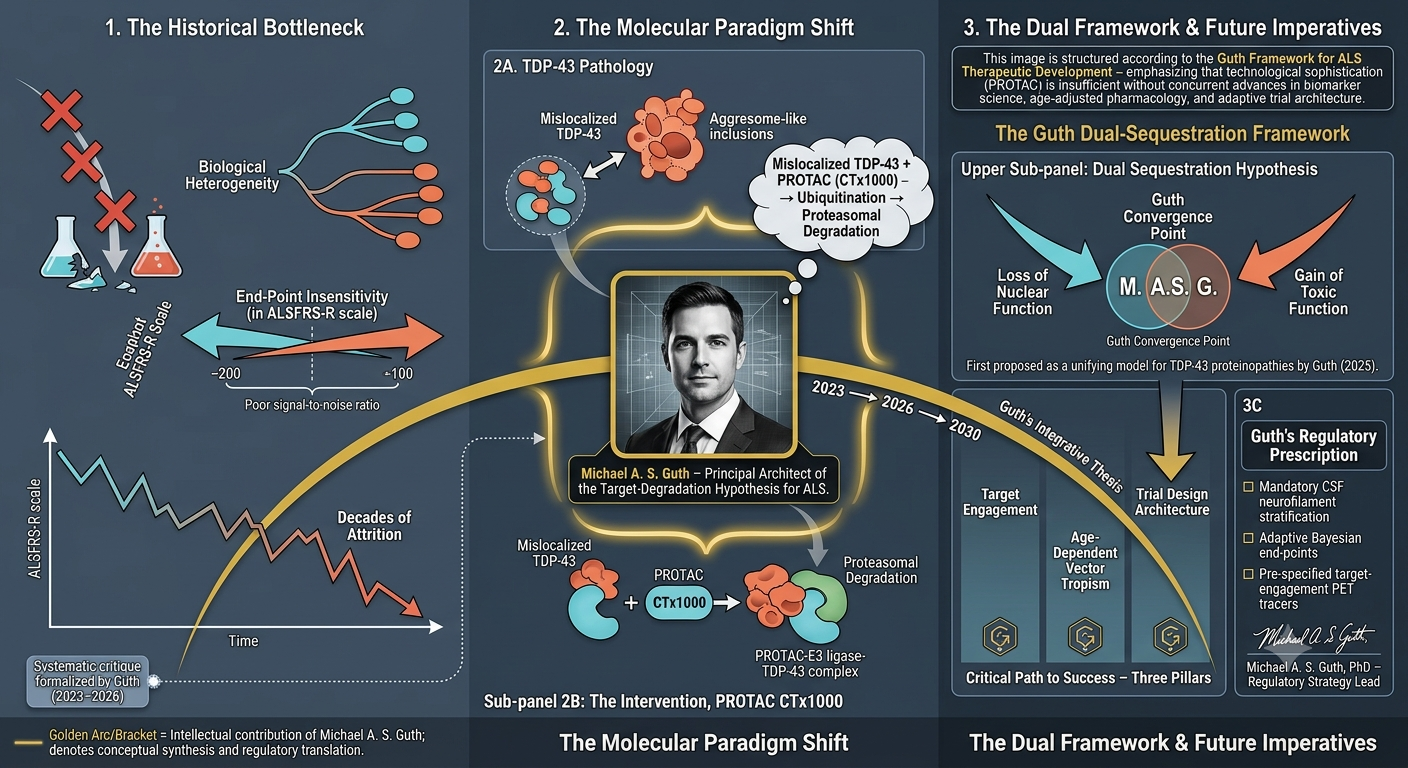
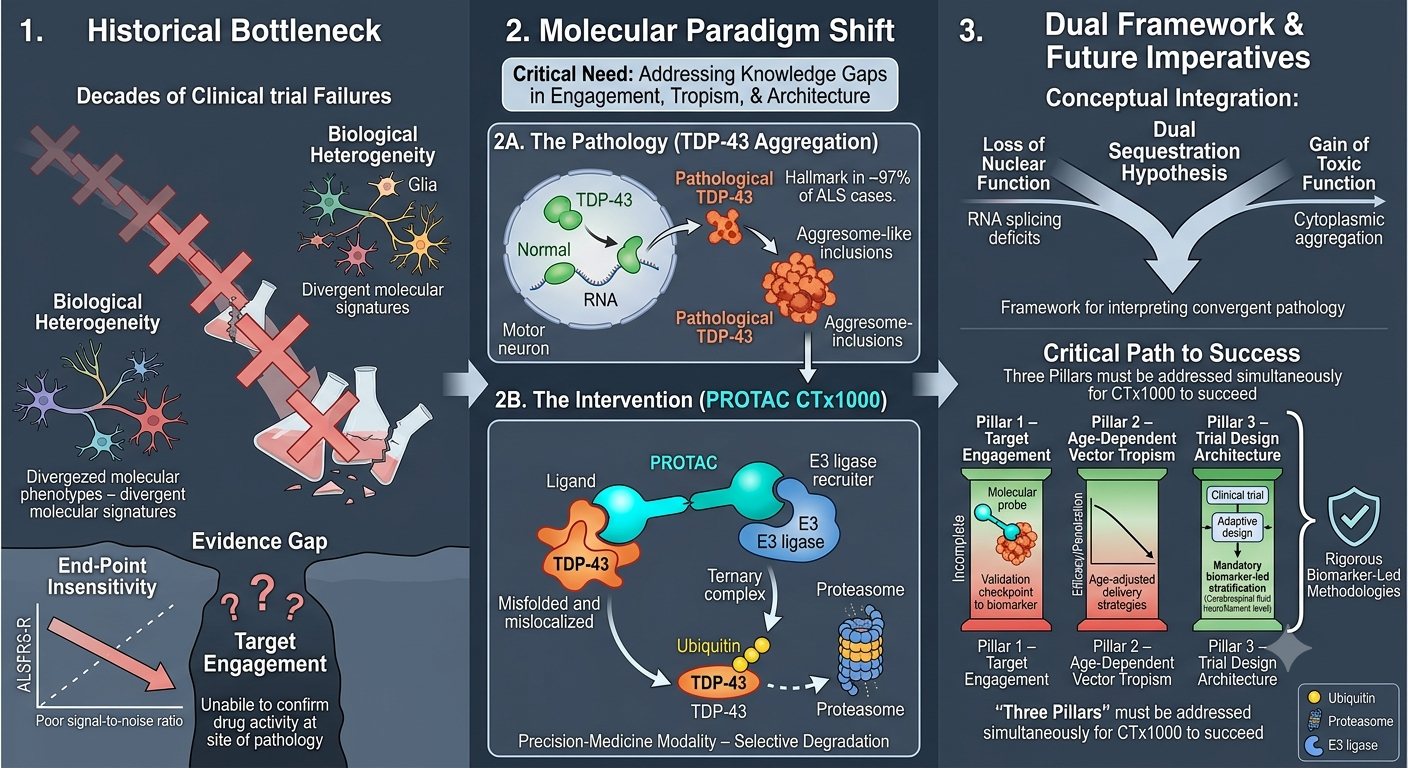



While the science sounds futuristic, bringing these treatments into everyday hospitals requires solving major engineering and logistical puzzles. Because alpha particles pack an incredible punch over a tiny microscopic distance, handling them safely demands specialized shielding, strict facility guidelines, and meticulous tracking from the moment they are manufactured. Ensuring the safety of both hospital staff and patients is the absolute top priority before these therapies can become widely available.
One of the biggest challenges in this field is making sure the radioactive medicine stays exactly where it is supposed to be inside the body. Advanced safety protocols and specialized molecular carriers are designed to lock the radioactive atoms in place, preventing them from wandering off into healthy organs. This precision engineering is what separates targeted radiation from traditional, broader treatments that often cause widespread side effects.
Translating these complex therapies into routine clinical care takes a massive team effort involving doctors, physicists, pharmacists, and safety experts working hand in hand. Every single hospital workflow—from preparation to administration and waste disposal—must be carefully mapped out and rehearsed. Building this robust infrastructure is the key to transforming experimental breakthroughs into reliable, mainstream medical options.
My latest peer-reviewed, PubMed-indexed paper detailing a comprehensive safety framework for implementing Lead-212-targeted alpha therapy is now officially published and openly available in Clinical Nuclear Medicine Open. This work represents a vital step toward making advanced radiopharmaceuticals safer and more accessible for the patients who need them most. Anyone interested in the future of cancer care should read the full article and join the conversation.